
Sowerby’s Beaked Whales (Mesoplodon bidens) in the Skagerrak and Adjacent Waters: Historical Records and Recent Post-Mortem Findings

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Historical Records
2.2. Stranding Events 2015–2020
2.3. Post-Mortem Examinations
2.4. Diet Analysis
3. Results
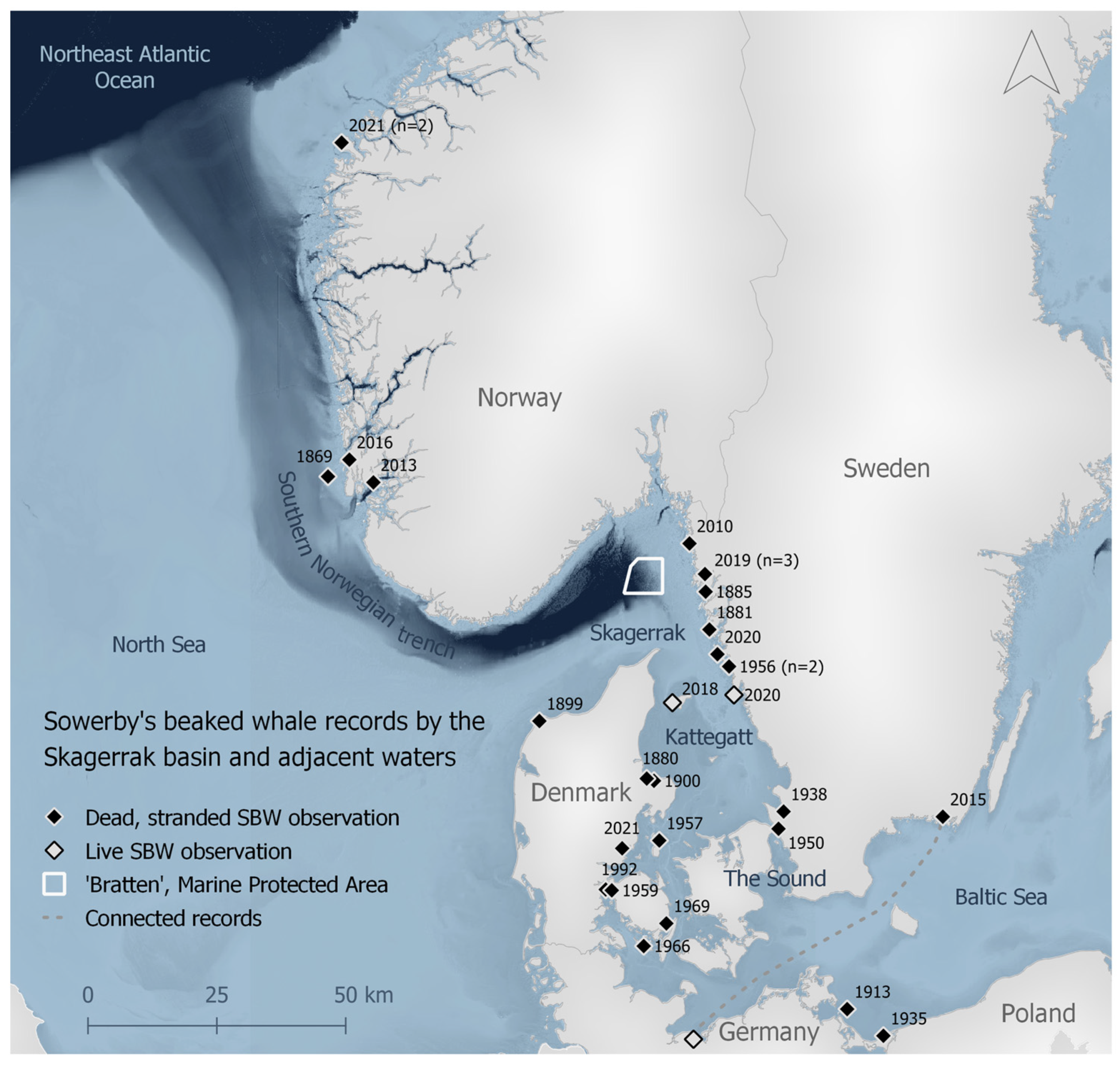
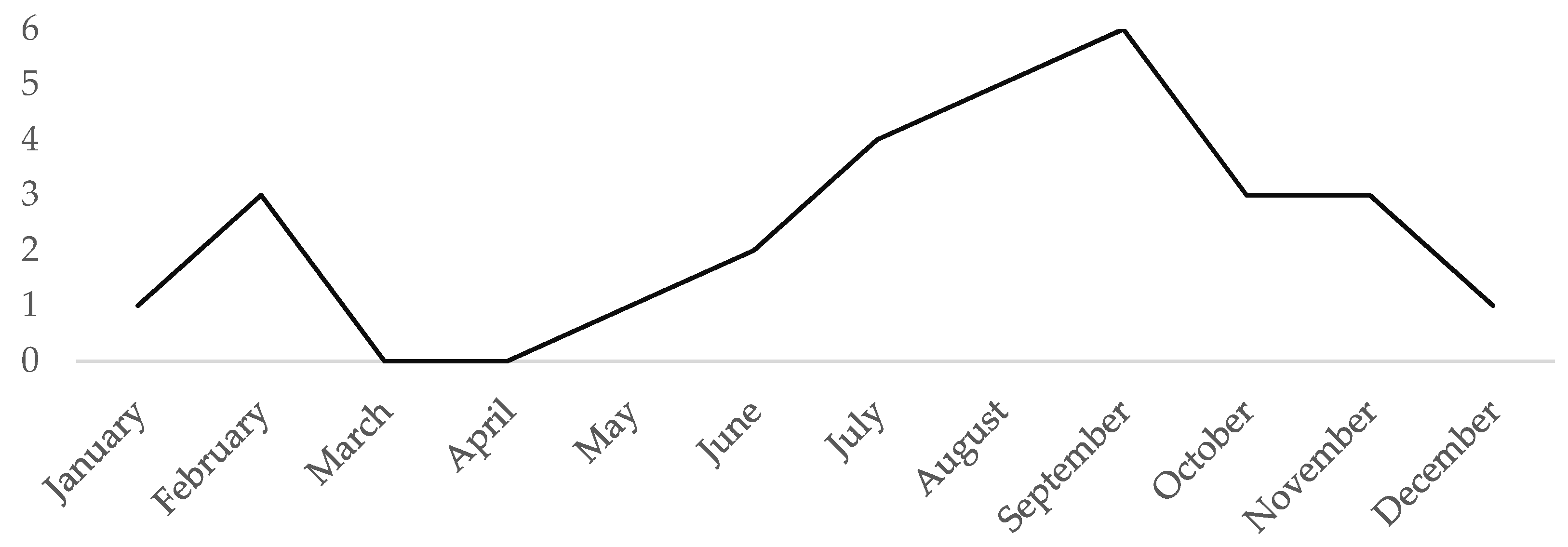
3.1. Historical Records
3.2. Description of Recent Stranding Events 2015–2020
3.2.1. October 2015
3.2.2. August 2019
3.2.3. July 2020
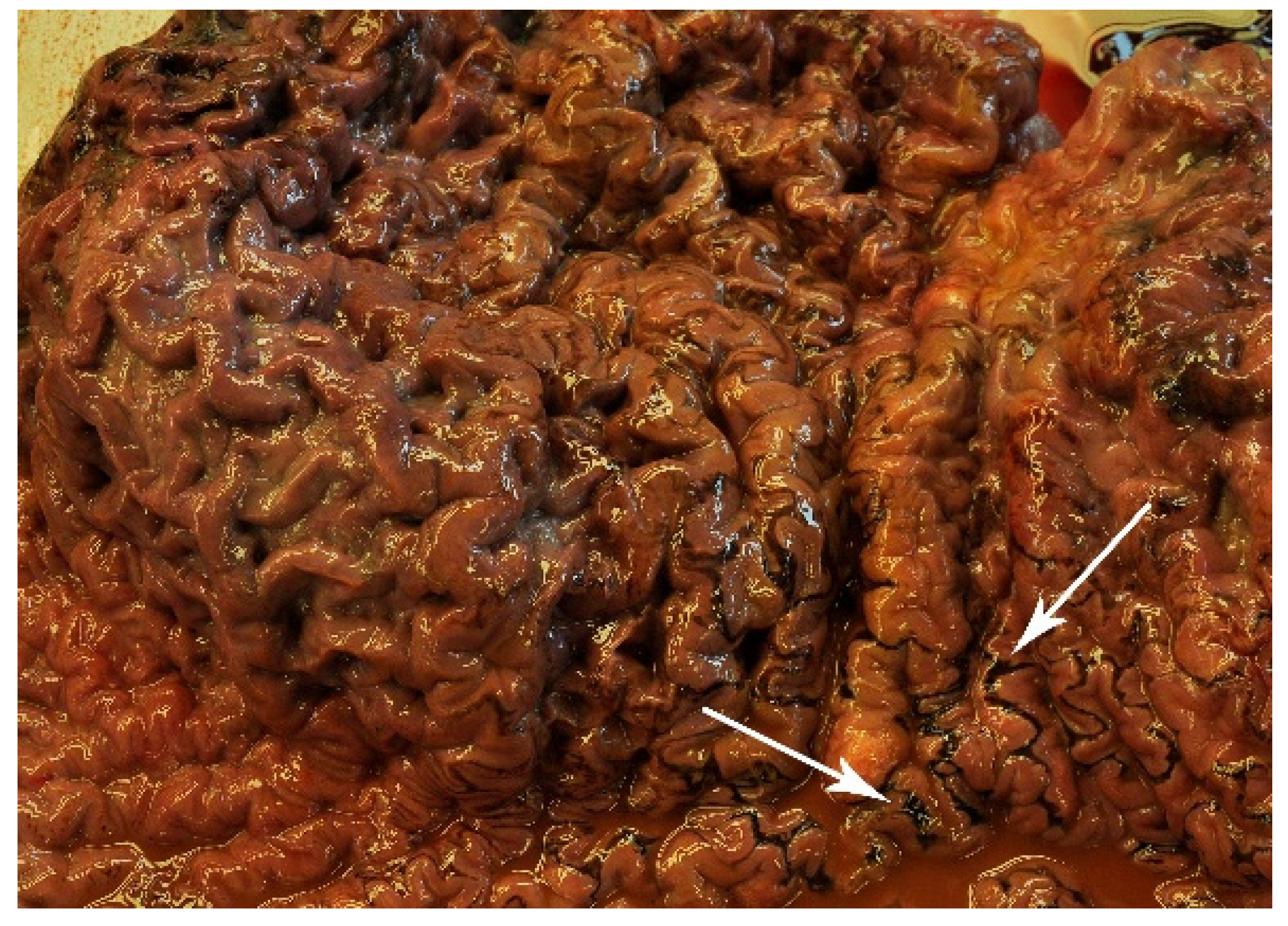
3.3. Necropsy Findings
3.3.1. Specimen 1, ID 15-VLT002234
3.3.2. Specimen 2, ID: 19-VLT001658
3.3.3. Specimen 3 ID: 19-VLT001659
3.3.4. Specimen 4, ID: NRM A2019/05472
3.3.5. Specimen 5, ID: 20-VLT001629
3.4. Diet Analysis
4. Discussion
4.1. Historical Records of SBWs
4.2. Necropsy Findings
4.3. Diet Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year | Month | Sex | Age | Length (m) | Locality | Country | Note |
|---|---|---|---|---|---|---|---|
| 1869 | June | Male | Adult | 4.40 | Nord Jan | Norway | |
| 1880 | February | Female | Adult | 4.32 | Hevringholm | Denmark | |
| 1881 | November | Male | Adult | 4.50 | Marstrand | Sweden | |
| 1885 | August | Male | Unavailable | 3.87 | Saltö, Lysekil | Sweden | Live stranded |
| 1899 | December | Male | Juvenile | 4.27 | Klitmöller | Denmark | |
| 1900 | October | Female | Adult | 5.02 | Hevringholm | Denmark | Pregnant |
| 1913 | July | Female | Juvenile | Unavailable | Griefswalde oie | Germany | Live stranded, killed |
| 1935 | June | Male | Adult | 4.50 | Wolin | Poland | Live stranded, killed |
| 1938 | September | Male | Adult | 4.50 | Vegeholm | Sweden | |
| 1950 | September | Male | Adult | 4.47 | Helsingborg | Sweden | |
| 1956 | September | Female | Juvenile | 3.20 | Askimfjorden | Sweden | Mass stranded |
| 1956 | September | Female | Adult | 4.75 | Askimfjorden | Sweden | Mass stranded |
| 1957 | November | Male | Adult | 5.00 | Stavsfjord | Denmark | |
| 1959 | September | Female | Adult | ~5 | Gamboerg Fjord | Denmark | |
| 1966 | September | Unavailable | Unavailable | ~4 | Marstal | Denmark | |
| 1969 | January | Prob. Female | Adult | ~5 | Lundeborg | Denmark | |
| 1992 | October | Female | Juvenile | 3.34 | Stenderup hage | Denmark | |
| 2010 | * June | Unavailable | Unavailable | Unavailable | Tjurpannans nature reserve | Sweden | Skeletal remains |
| 2013 | November | Female | Unavailable | Unavailable | Rogaland | Norway | |
| 2015 | October | Female | Juvenile | 3.42 | Karlskrona | Sweden | |
| 2016 | February | Unavailable | Unavailable | Unavailable | Rogaland | Norway | |
| 2018 | February | Male | Adult | ~5 | Læsø | Denmark | Live stranded, helped out |
| 2019 | August | Male | Juvenile | 4.55 | Fiskebäckskil | Sweden | Live, mass stranded |
| 2019 | August | Male | Juvenile | 4.24 | Fiskebäckskil | Sweden | Live, mass stranded |
| 2019 | August | Male | Juvenile | 4.47 | Fiskebäckskil | Sweden | Live, mass stranded |
| 2020 | May | Unavailable | Unavailable | ~4 | Tistlarna | Sweden | Alive |
| 2020 | July | Male | Juvenile | 3.85 | Öckerö | Sweden | Euthanized |
| 2021 | July | Unavailable | Unavailable | Unavailable | Ervik | Norway | Live stranded |
| 2021 | July | Unavailable | Calf | Unavailable | Ervik | Norway | |
| 2021 | August | Female | Juvenile | 3.62 | Horsens fjord | Denmark |
References
- Macleod, C.D. Review of the distribution of Mesoplodon species (order Cetacea, family Ziphiidae) in the North Atlantic. Mammal Rev. 2000, 30, 1–8. [Google Scholar] [CrossRef]
- International Union for Conservation of Nature (IUCN). Sowerby’s Beaked Whale. Available online: https://www.iucnredlist.org/species/13241/50363686 (accessed on 24 February 2022).
- Smith, K.J.; Mead, J.G.; Peterson, M.J. Specimens of opportunity provide vital information for research and conservation regarding elusive whale species. Environ. Conserv. 2021, 48, 1–9. [Google Scholar] [CrossRef]
- Smith, K.J.; Trueman, C.N.; France, C.A.M.; Sparks, J.P.; Brownlow, A.C.; Dähne, M.; Davison, N.J.; Guðmundsson, G.; Khidas, K.; Kitchener, A.C.; et al. Stable isotope analysis of specimens of opportunity reveals ocean-scale site fidelity in an elusive whale species. Front. Conserv. Sci. 2021, 2, 13. [Google Scholar] [CrossRef]
- Carlström, J.; Denkinger, J.; Feddersen, P.; Øien, N. Record of a new northern range of Sowerby’s beaked whale (Mesoplodon bidens). Polar Biol. 1997, 17, 459–461. [Google Scholar] [CrossRef]
- Reiner, F. First record of Sowerby’s beaked whale from Azores. Sci. Rep. Whales Res. Inst. 1986, 37, 103–170. [Google Scholar]
- Bachara, W.; Barreiros, J.P. Stranding of Four Sowerby’s Beaked Whales (Mesoplodon bidens) in Terceira, Azores, Unpublished Report to Cetal Fauna WB2016/3. 2016.
- Bittau, L.; Leone, M.; Gannier, A.; Gannier, A.; Manconi, R. Two live sightings of Sowerby’s beaked whale (Mesoplodon bidens) from the western Mediterranean (Tyrrhenian Sea). Marine Biological Association of the United Kingdom. J. Mar. Biol. Assoc. United Kingd. 2018, 98, 1003–1009. [Google Scholar] [CrossRef]
- Martin, V.; Tejedor, M.; Perez-Gil, M.; Dalebout, M.L.; Arbelo, M.; Fernandez, A. A Sowerby’s beaked whale (Mesoplodon bidens) stranded in the Canary Islands: The most southern record in the eastern North Atlantic. Aquat. Mamm. 2011, 37, 512–519. [Google Scholar] [CrossRef]
- Rogan, E.; Cañadas, A.; MacLeod, K.; Begoña Santos, M.; Mikkelsen, B.; Uriarte, A.; Van Canneyt, O.; Vázquez, J.A.; Hammond, P.S. Distribution, abundance and habitat use of deep diving cetaceans in the North-East Atlantic. Deep Sea Res. Part II Top. Stud. Oceanogr. 2017, 141, 8–19. [Google Scholar] [CrossRef] [Green Version]
- Berrow, S.; Meade, R.; Marrinan, M.; McKeogh, E.; O’Brien, J. First confirmed sighting of Sowerby’s beaked whale (Mesoplodon bidens (Sowerby, 1804)) with calves in the Northeast Atlantic. Mar. Biodivers. Rec. 2018, 11, 20. [Google Scholar] [CrossRef]
- Robbins, J.R.; Bell, E.; Potts, J.; Babey, L.; Marley, S.A. Likely year-round presence of beaked whales in the Bay of Biscay. Hydrobiologia 2022, 849, 2225–2239. [Google Scholar] [CrossRef]
- Global Biodiversity Information Facility (GBIF). Available online: https://www.gbif.org/occurrence/download/0154375-210914110416597 (accessed on 24 February 2022).
- MacLeod, C.D.; Pierce, G.J.; Santos, M.B. Geographic and temporal variations in strandings of beaked whales (Ziphiidae) on the coasts of the UK and the Republic of Ireland from 1800–2002. J. Cetacean Res. Manag. 2004, 6, 79–86. [Google Scholar]
- MacLeod, C.D.; Perrin, W.F.; Pitman, R.; Barlow, J.; Ballance, L.; D’Amico, A.; Gerrodette, T.; Joyce, G.; Mullin, K.D.; Palka, D.L.; et al. Known and inferred distributions of beaked whale species (Cetacea: Ziphiidae). J. Cetacean Res. Manag. 2005, 7, 271–286. [Google Scholar]
- Smeenk, C. Strandings of sperm whales Physeter macrocephalus in the North Sea: History and patterns. Bull. Inst. R. Sci. Nat. Belg. Biol. 1997, 67, 15–28. [Google Scholar]
- Brownlow, A.; Davison, N.; ten Doeschate, M. Scottish Marine Animal Stranding Scheme (SMASS); Annual Report; Scotland’s Rural College (SRUC): Aberdeen, UK, 2018. [Google Scholar]
- Davison, N.; ten Doeschate, M.; Brownlow, A. Scottish Marine Animal Stranding Scheme (SMASS); Annual Report; Scotland’s Rural College (SRUC): Aberdeen, UK, 2019. [Google Scholar]
- Sierra, E.; Ramírez, T.; Bernaldo de Quirós, Y.; Arregui, M.; Mompeó, B.; Rivero, M.A.; Fernández, A. Pulmonary and Systemic Skeletal Muscle Embolism in a Beaked Whale with a Massive Trauma of Unknown Aetiology. Animals 2022, 12, 508. [Google Scholar] [CrossRef] [PubMed]
- Davison, N.J.; Brownlow, A.; Doeschate, M.T.; Dale, E.J.; Foster, G.; Muchowski, J.; Perrett, L.L.; Rocchi, M.; Whatmore, A.M.; Dagleish, M.P. Neurobrucellosis due to Brucella ceti ST26 in Three Sowerby’s Beaked Whales (Mesoplodon bidens). J. Comp. Pathol. 2021, 182, 1–8. [Google Scholar] [CrossRef]
- Fernández, A.; Edwards, J.F.; Rodríguez, F.; Espinosa de los Monteros, A.; Herráez, P.; Castro, P.; Jaber, J.R.; Martín, V.; Arbelo, M. “Gas and fat embolic syndrome” involving a mass stranding of beaked whales (family Ziphiidae) exposed to anthropogenic sonar signals. Vet. Pathol. 2005, 42, 446–457. [Google Scholar] [CrossRef]
- Cox, T.M.; Ragen, T.J.; Read, A.J.; Vos, E.; Baird, R.W.; Balcomb, K.; Barlow, J.; Caldwell, J.; Cranford, T.; Crum, L.; et al. Understanding the impacts of anthropogenic sound on beaked whales. J. Cetacean Res. Manag. 2006, 7, 177–187. [Google Scholar]
- Filadelfo, R.; Mintz, J.; Michlovich, E.; D’Amico, A.; Tyack, P.L.; Ketten, D.R. Correlating military sonar use with beaked whale mass strandings: What do the historical data show? Aquat. Mamm. 2009, 35, 435–444. [Google Scholar] [CrossRef] [Green Version]
- Dolman, S.J.; Berrow, S.D.; Brownlow, A.; Deaville, R.; Evans, P.G.H.; Fernandez, A.; Gordon, J.C.D.; Haelters, J.; Ijsseldijk, L.L.; Miller, P.; et al. Overcoming Challenges to Protect Beaked Whales in the Northeast Atlantic; ASCOBANS Intersessional Working Group Report, ASCOBANS: 26th Meeting of the Advisory Committee; ASCOBANS: Bonn, Germany, 2021; Available online: https://www.ascobans.org/sites/default/files/document/ascobans_ac26_doc4.3_iwg-beaked-whales-report.pdf (accessed on 17 March 2022).
- MacLeod, C.D.; Santos, M.B.; Pierce, G.J. Review of data on diets of beaked whales: Evidence of niche separation and geographic segregation. J. Mar. Biol. Assoc. United Kingd. 2003, 83, 651–665. [Google Scholar] [CrossRef]
- Pereira, J.N.; Neves, V.C.; Prieto, R.; Silva, M.A.; Cascão, I.; Oliveira, C.; Cruz, M.J.; Medeiros, J.V.; Barreiros, J.P.; Porteiro, F.M.; et al. Diet of mid-Atlantic Sowerby’s beaked whales Mesoplondon bidens. Deep. Sea Res. Part I Oceanogr. Res. Pap. 2011, 58, 1084–1090. [Google Scholar] [CrossRef] [Green Version]
- Wenzel, F.W.; Polloni, P.T.; Craddock, J.E.; Gannon, D.P.; Nicolas, J.R.; Read, A.J.; Rosel, P.E. Food habits of Sowerby’s beaked whales (Mesoplodon bidens) taken in the pelagic drift gillnet fishery of the western North Atlantic. Fish. Bull. 2013, 111, 381–389. [Google Scholar] [CrossRef] [Green Version]
- Kinze, C.C. Ny stranding af almindelig næbhval (Mesoplodon bidens) og oversigt over hidtidige danske fund. Flora Fauna 1993, 99, 99–104. [Google Scholar]
- Kinze, C.C.; Waller, G.N.H. Nineteenth-century records of Sowerby’s beaked whale (Mesoplodon bidens): Addendum, Danish records. Arch. Nat. Hist. 2015, 42, 357–358. [Google Scholar] [CrossRef]
- Kinze, C.C.; Thøstesen, C.B.; Olsen, M.T. Cetacean stranding records along the Danish coastline: Records for the period 2008–2017 and a comparative review. Lutra 2018, 61, 87–106. [Google Scholar]
- The Norwegian Biodiversity Information Centre. Norwegian Species Observation Service. Available online: https://ipt.artsdatabanken.no/resource?r=speciesobservationsservice2&request_locale=en (accessed on 21 December 2021).
- Waller, G.N.H. A review of nineteenth-century records of Sowerby’s beaked whale (Mesoplodon bidens). Arch. Nat. Hist. 2014, 41, 338–356. [Google Scholar] [CrossRef]
- Geraci, J.R.; Lounsbury, V.R. Marine Mammals Ashore: A Field Guide for Strandings, 2nd ed.; National Aquarium in Baltimore: Baltimore, MD, USA, 2005; 305p. [Google Scholar]
- Valar.se Om Valar i Svenska Vatten. Available online: http://www.valar.se (accessed on 3 March 2022).
- Kuiken, T.; Hartman, M.G. Cetacean Pathology: Dissection Techniques and Tissue Sampling. In Proceedings of the First European Cetacean Society Workshop, Leiden, The Netherlands, 13–14 September 1991. [Google Scholar]
- IJsseldijk, L.L.; Brownlow, A.C.; Mazzariol, S. European Best Practice on Cetacean Post-mortem Investigation and Tissue Sampling; Joint ACCOBAMS and ASCOBANS Document. 2019. Available online: https://osf.io/zh4ra/ (accessed on 3 March 2022).
- HELCOM. Administrative Boundaries. Available online: https://metadata.helcom.fi/geonetwork/srv/eng/catalog.search#/metadata/b58b0b56-1a98-41d9-9bf4-5dc76f5069d3 (accessed on 28 December 2021).
- EMODnet Digital Bathymetry 2020. Available online: https://sextant.ifremer.fr/record/bb6a87dd-e579-4036-abe1-e649cea9881a/ (accessed on 14 November 2021).
- Lantmäteriet (MPA, OSPAR). Available online: https://www.geodata.se/geodataportalen/ (accessed on 12 January 2022).
- Bancroft, J.D.; Cook, H.C. Manual of Histological Techniques; Churchill Livingstone: Edinburgh, UK, 1984. [Google Scholar]
- Siebert, U.; Wünschmann, A.; Weiss, R.; Frank, H.; Benke, H.; Frese, K. Post mortem findings in harbour porpoises (Phocoena phocoena) from the German North and Baltic Sea. J. Comp. Pathol. 2001, 124, 102–114. [Google Scholar] [CrossRef] [PubMed]
- Lehnert, K.; Raga, J.A.; Siebert, U. Macroparasites in stranded and bycaught harbour porpoises from German and Norwegian waters. Dis. Aquat. Org. 2005, 64, 265–269. [Google Scholar] [CrossRef] [PubMed]
- Otoliths of North Sea Fish 1.0. Available online: https://otoliths-northsea.linnaeus.naturalis.nl/ (accessed on 3 February 2022).
- Miya, M.; Sato, Y.; Fukunaga, T.; Sado, T.; Poulsen, J.Y.; Sato, K.; Minamoto, T.; Yamamoto, S.; Yamanaka, H.; Araki, H.; et al. MiFish, a set of universal PCR primers for metabarcoding environmental DNA from fishes: Detection of more than 230 subtropical marine species. R. Soc. Open Sci. 2015, 2, 150088. [Google Scholar] [CrossRef] [Green Version]
- Martin, M. Cutadapt Removes Adapter Sequences from High-Throughput Sequencing Reads. EMBnet J. 2011, 17, 10–12. [Google Scholar] [CrossRef]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. Dada2: High-Resolution Sample Inference from Illumina Amplicon Data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [Green Version]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Mead, J.G. Stomach anatomy and use in defining systemic relationships of the Cetacean family Ziphiidae (beaked whales). Anat. Rec. 2007, 290, 581–595. [Google Scholar] [CrossRef] [PubMed]
- Evans, P.G.H.; Hammond, P.S. Monitoring cetaceans in European waters. Mammal Rev. 2004, 34, 131–156. [Google Scholar] [CrossRef]
- Peltier, H.; Baagøe, H.J.; Camphuysen, K.C.J.; Czeck, R.; Dabin, W.; Daniel, P.; Deaville, R.; Haelters, J.; Jauniaux, T.; Jensen, L.F.; et al. The stranding anomaly as population indicator: The case of harbour porpoise Phocoena phocoena in North-Western Europe. PLoS ONE 2013, 8, e62180. [Google Scholar] [CrossRef] [Green Version]
- Whitehead, H. Trends in cetacean abundance in the Gully submarine canyon, 1988–2011, highlight a 21% per year increase in Sowerby’s beaked whales (Mesoplodon bidens). Can. J. Zool. 2013, 91, 141–148. [Google Scholar] [CrossRef] [Green Version]
- Florén, K.; Nicolopoulos, A. Revmiljöer i Skageraks Utsjö—En Litteraturstudie och Sammanställning av Inventeringsdata. AquaBiota Report. 2015. Available online: https://www.aquabiota.se/wp-content/uploads/Floren_Nikolopoulos_Revmiljoer-i-Skagerak_AqB_Notes_2015_02.pdf (accessed on 15 March 2022). AquaBiota Report.
- Kilnäs, M. Förslag till Fiskeregleringar i Bratten. Report from the Project Hav Möter Land. 2013. Available online: https://interreg-oks.eu/download/18.2f79a9231506ca113761737a/1472022928091/Hav%20mxder%20land%20Bratten.pdf (accessed on 15 March 2022).
- Visser, F.; Oudejans, M.G.; Keller, O.A.; Madsen, P.T.; Johnson, M. Sowerby’s beaked whale biosonar and movement strategy indicate deep-sea foraging niche differentiation in mesoplodont whales. J. Exp. Biol. 2022, 225, jeb243728. [Google Scholar]
- Barile, C.; Berrow, S.; O’Brien, J. Oceanographic Drivers of Cuvier’s (Ziphius cavirostris) and Sowerby’s (Mesoplodon bidens) Beaked Whales Acoustic Occurrence along the Irish Shelf Edge. J. Mar. Sci. Eng. 2021, 9, 1081. [Google Scholar] [CrossRef]
- Kowarski, K.; Delarue, J.; Martin, B.; O’Brien, J.; Meade, R.; Cadhla, O.Ó.; Berrow, S. Signals from the deep: Spatial and temporal acoustic occurrence of beaked whales off western Ireland. PLoS ONE 2018, 13, e0199431. [Google Scholar] [CrossRef]
- New, L.F.; Moretti, D.J.; Hooker, S.K.; Costa, D.P.; Simmons, S.E. Using energetic models to investigate the survival and reproduction of beaked whales (family Ziphiidae). PLoS ONE 2013, 8, e68725. [Google Scholar] [CrossRef] [Green Version]
- Murie, D.J.; Lavigne, D.M. Interpretation of otoliths in stomach content analyses of phocid seals: Quantifying fish consumption. Can. J. Zool. 1986, 64, 1152–1157. [Google Scholar] [CrossRef]
- Wijnsma, G.; Pierce, G.J.; Santos, M.B. Assessment of errors in cetacean diet analysis: In vitro digestion of otoliths. J. Mar. Biol. Assoc. UK 1999, 79, 573–575. [Google Scholar] [CrossRef]
- Heyning, J.E. Suction feeding in beaked whales: Morphological and observational evidence. Nat. Hist. Mus. Los Angeles Cty. Contrib. Sci. 1996, 464, 1–12. [Google Scholar] [CrossRef]









| Gene Fragment | Primer Name | Primer F | Primer R |
|---|---|---|---|
| 16S long | 16sPreyF | CGTGCRAAGGTAGCG | GTCGCCCCAACCRAAG |
| 16S short | 16sFiskF | GAAGACCCTWTGGAGCTTWAG | GTCGCCCCAACCRAAG |
| 12S | MiFish | AAACTCGTGCCAGCCACC | GGGTATCTAATCCCAGTTTG |
| Specimen 1 | Specimen 2 | Specimen 3 | Specimen 4 | Specimen 5 | |
|---|---|---|---|---|---|
| Date found dead | 2015-10-23 | 2019-08-07/08 | 2019-08-07/08 | 2019-08-05 | 2020-07-11 |
| Location | Karlshamn (Baltic Sea) | Fiskebäckskil (Skagerrak) | Fiskebäckskil (Skagerrak) | Fiskebäckskil (Skagerrak) | Öckerö (Skagerrak/Kattegatt) |
| Sex | Female | Male | Male | Male | Male |
| Age class | Sub-adult | Sub-adult | Sub-adult | Sub-adult | Sub-adult |
| Length | 3.42 m | 4.55 m | 4.24 m | 4.47 m | 3.85 m |
| Weight | 340 kg | 826 kg | 730 kg | N/A | N/A |
| DCC1–5 | 3 | 4 | 3–4 | N/E | 3–4 |
| Cause of death | Emaciated | Live stranding, undetermined cause | Live stranding, undetermined cause | Live stranding, undetermined cause | Euthanized, traumatic wounds |
| Other findings | Pock marks, gastric ulcerations | Pock marks | Focal osteomyelitis (maxilla), pock marks, gastric ulceration, tooth rake marks | Osteomyelitis (vertebrae) | Possible pock marks |
| Parasites | 30–40 nematodes in stomach | Few trematodes in a bile duct |
| Prey List of Lower Taxa | Specimen 2 | Specimen 3 | Specimen 5 | Total | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| n | N | %N | n | N | %N | n | N | %N | n | N | %N | |
| Gadidae | ||||||||||||
| Gadidae spp. | 6 | 3 | 13.63 | 115 | 58 | 25.55 | 121 | 61 | 24.60 | |||
| Trisopterus sp. | 37 | 19 | 86.36 | 1 | 1 | 100.00 | 234 | 118 | 51.98 | 272 | 136 | 54.84 |
| Pleuronectidae | ||||||||||||
| Pleuronectidae spp. | 76 | 38 | 16.74 | 76 | 38 | 15.32 | ||||||
| Gobiidae | ||||||||||||
| Gobiidae spp. | 23 | 12 | 5.29 | 23 | 12 | 4.84 | ||||||
| Ostreidae | ||||||||||||
| Ostridae sp. | 1 | 1 | 0.44 | 1 | 1 | 0.40 | ||||||
| Total | 43 | 22 | - | 1 | 100 | 449 | 227 | - | 493 | 248 | - | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stavenow, J.; Roos, A.M.; Ågren, E.O.; Kinze, C.; Englund, W.F.; Neimanis, A. Sowerby’s Beaked Whales (Mesoplodon bidens) in the Skagerrak and Adjacent Waters: Historical Records and Recent Post-Mortem Findings. Oceans 2022, 3, 250-267. https://doi.org/10.3390/oceans3030018
Stavenow J, Roos AM, Ågren EO, Kinze C, Englund WF, Neimanis A. Sowerby’s Beaked Whales (Mesoplodon bidens) in the Skagerrak and Adjacent Waters: Historical Records and Recent Post-Mortem Findings. Oceans. 2022; 3(3):250-267. https://doi.org/10.3390/oceans3030018
Chicago/Turabian StyleStavenow, Jasmine, Anna Maria Roos, Erik Olof Ågren, Carl Kinze, William F. Englund, and Aleksija Neimanis. 2022. "Sowerby’s Beaked Whales (Mesoplodon bidens) in the Skagerrak and Adjacent Waters: Historical Records and Recent Post-Mortem Findings" Oceans 3, no. 3: 250-267. https://doi.org/10.3390/oceans3030018